

️ Vai trò y học của mạt
Mạt (liên họ mạt Gamasoidea) thuộc bộ Acarina và phụ bộ Mesostigmata. Đến nay, người ta đã thống kê được hơn 2.000 loài, thuộc 300 giống và phân giống mạt thuộc 20 họ, phân bố khắp địa cầu. Chúng là những sinh vật có kích thước nhỏ hơn 1 mm. Mạt có thể phân chia dựa vào hoạt động ký sinh, có loài ký sinh đốt máu, sống tự do hoặc vừa ký sinh vừa sống tự do hoặc ký sinh trong đường hô hấp của các động vật khác. Đặc biệt nhóm ký sinh đốt máu được quan tâm vì chúng có thể là véc tơ lan truyền bệnh. Quan tâm chính đến các loài mạt có vai trò lan truyền bệnh tập trung vào các họ Dermanyssidae, Macronyssidae và Laelapidae.
LAN TRUYỀN VI RÚT
Các vi rút gây bệnh ở động vật như fowlpox và Newcastle cho thấy có nhiều mối liên quan đến các loài mạt (Shirinov và cộng sự, 1972; Arzey, 1990). Nhóm hantavirus là một ví dụ về một trong những bệnh nguy hiểm được lan truyền từ động vật sang người và nhóm vi rút này đã được tìm thấy trên các loài mạt (Wu và cộng sự, 1998; Zhuge và cộng sự, 1998; Iakimenko và cộng sự, 2000; Zhang và cộng sự, 2001; Zhang và cộng sự, 2002).
Bảng 1. Các loài mạt có khả năng lan truyền vi rút
.png)
.png)
(A. casalis = Androlaelaps casalis, A. fahrenholzi = Androlaelaps fahrenholzi, A. glasgowi = Androlaelaps glasgowi, B. sylviarum = Bdellonyssus sylviarum, D. americanus = Dermanyssus americanus, D. hirundinis = Dermanyssus hirundinis, E. shanghaiensis = Eulaelaps shanghaiensis, E. stabularis = Eulaelaps stabularis, H. ambulans = Haemogamasus ambulans, H. glasgowi = haemolaelaps glasgowi, A. hirsutus = Hae- mogamasus hirsutus, H. isabellinus = Hirstionyssus isabellinus, H. nidi = Haemogamasus nidi, H. mandschuicus = Haemolaelaps mand- schuicus, H. serdjukovae = Haemogamasus serdjukovae, H. talpae = Hirstionyssus talpae, L. agilis = Laelaps agilis, L. clethrionomydis = Lae- laps clethrionomydis, O. bacoti = Ornithonyssus bacoti, O. bursa = Ornithonyssus bursa, O. sylviarum = Ornithonyssus sylviarum).
Hơn nữa, đối với một số loài, lan truyền vi rút theo cơ chế Trans-tadial và Trans-ovarian đã được chứng minh, và có khả năng lan truyền sang các loài gặm nhấm qua vết đốt (Meng và cộng sự, 1990; Đồng, 1991). Tương tự như vậy, các vi rút gây viêm não Saint-Louis đã được phân lập từ các loài mạt này nhưng không có bằng chứng các loài này có khả năng lan truyền hay là vật chứa vi rút gây viêm não Saint-Louis (Reeves và cộng sự, 1955;.. Chamberlain và cộng sự, 1957). Các loại vi rút khác, chẳng hạn như vi rút Tây sông Nile, các vi rút gây viêm não do ve truyền và nhóm togaviruses cũng đã được phân lập từ các loài mạt này: trong một số trường hợp, mạt có khả năng là véc tơ lan truyền bệnh (Zemskaya & Pchel- kina, 1962; Gil'Manova và cộng sự, 1964; Chernykh và cộng sự, 1974; Zemskaya & Pchelkina, 1974a; Zemskaya & Pchelkina, 1974b; Kornilova và cộng sự, 1975;. Shaiman & Tarasevich, 1976; Wegner, 1976; Naumov & Gutova, 1984; Kocianova & Kozuch năm 1988;. Iakimenko và cộng sự, 1991; Durden & Turell năm 1993;. Kocianova và cộng sự, 1993;. Korenberg và cộng sự, 1993). Nhóm togaviruses, gây viêm não ngựa phương Tây, phương Đông và Venezuela cần phải được nghiên cứu kỹ hơn.
Viêm não ngựa phương Đông và phương Tây là những mô hình bệnh tật ở Mỹ cũng như Venezuela, nó gây ra rối loại thần kinh nghiêm trọng ở cả người và ngựa. Bản chất của các rối loạn có thể là do viêm não hoặc viêm cột sống. Lan truyền xảy ra hầu như là do muỗi và tồn tại vĩnh viễn trong vật chủ (các loài chim và động vật gặm nhấm). Các loài mạt được cho là có khả năng lan truyền các bệnh viêm não này. Để đánh giá vai trò lan truyền các vi rút này của mạt, nhóm nghiên cứu đã tiến hành thí nghiệm bằng cách sử dụng phương pháp sau đây: cho các loài mạt đốt vật chủ bị nhiễm và sau đó kiểm tra sự hiện hiện của tác nhân gây bệnh ở mạt. Nếu mạt bị nhiễm bệnh, tiết tục cho mạt đốt vật chủ không bị nhiễm như thế có thể đánh gia khả năng lan truyền vi rút. Thế hệ sau của loài mạt bị nhiễm này sau đó cũng được phân tích để đánh giá khả năng lan truyền theo cơ chế Trans-ovarian.
Bảng 2. Các loài mạt có khả năng lan truyền vi rút viêm não ngựa
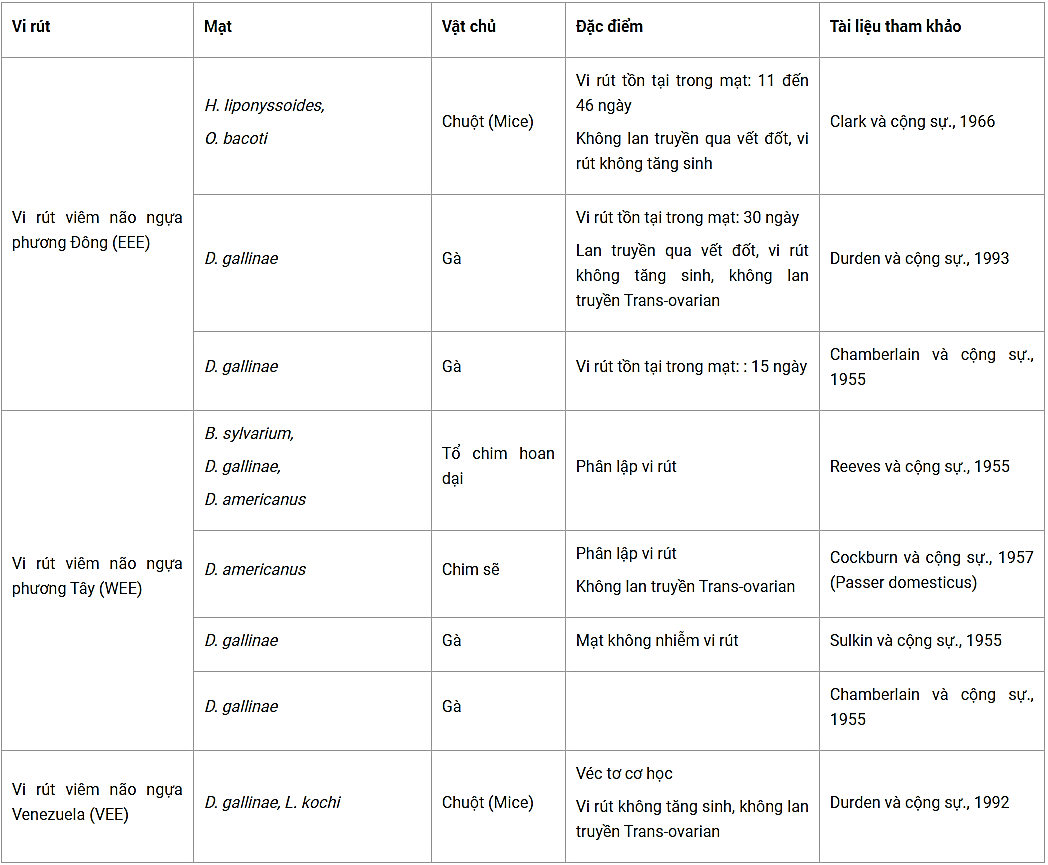
(B. sylviarum = Bdellonyssus sylviarum, D. americanus = Der- manyssus americanus, H. liponyssoides = Haemogamasus liponyssoides, L. kochi = Laelaps kochi, O. bacoti = Ornithonyssus bacoti).
Đối với các vi rút gây viêm não ngựa phương Đông (EEE), Clark và cộng sự. (1966) đã chứng minh rằng cả hai loài mạt Haemogamasus liponyssoides và Ornithonyssus bacoti có thể đưa vào cơ thể chúng một số lượng lớn vi rút khi chúng đốt máu chuột bị nhiễm. Tùy thuộc vào nhiệt độ, vi rút này có thể tồn tại lên đến 46 ngày trong cơ thể mạt nhưng vi rút không tăng sinh và không quan sát được khả năng lan truyền vi rút qua các vết đốt. Tuy nhiên, Chamberlain & Sikes (1955); Durden và cộng sự (1993) cho thấy rằng loài Dermanyssus gallinae đốt gà con bị nhiễm và mang vi rút ít nhất một tháng mà vi rút này không được tăng sinh và không lan truyền vi rút cho thế hệ sau. Mặc dù loài mạt này có thể lan truyền vi rút cho các con gà khác qua vết đốt khi chúng đốt máu. Thời gian ủ bệnh tối đa là 26 ngày trong cả hai trường hợp. Các xét nghiệm tìm vi rút từ phân của mạt đều âm tính.
Trong trường hợp của vi rút gây viêm não ngựa phương Tây (WEE), có 09 chủng vi rút được phân lập từ loài mạt Bdellonyssus sylviarum và Dermanyssus americanus giữa năm 1946 và 1949, hai loài mạt này được thu từ tổ chim hoang ở California (Reeves và cộng sự., 1955). Trong một trường hợp tương tự WEE được phân lập từ một người đàn ông ở Colorado, sau đó người ta phát hiện con chim sẻ ở trang trại của bệnh nhân bị loài mạt D. americanus ký sinh và ở trong con mạt này mang vi rút WWE (Cockburn và cộng sự., 1957). Tất cả các nghiên cứu trong phòng thí nghiệm để xác định vai trò lan truyền vi rút của mạt so với các kết qua trước đây có sự khác biệt rất lớn. Reeves và cộng sự (1955); Sulkin và cộng sự (1955) đã không thành công trong việc chứng minh khả năng lan truyền vi rút của hai loài mạt D. gallinae và B. sylviarum từ gà nhiễm bệnh và chim khỏe mạnh. Tuy nhiên, Cockburn và cộng sự (1957) nuôi loài mạt D. gallinae trên gà bị nhiễm vi rút, nhưng không thể chứng minh khả năng lan truyền sang các con chim khỏe mạnh khác hay lan truyền Trans-ovarian. Cuối cùng, Chamberlain và Sikes (1955) tiến hành thí nghiệm là gây nhiễm cho gà khỏe mạnh bằng cách cho D. gallinae chứa vi rút đốt máu, thời gian ủ bệnh là 13 ngày.
Đối với các vi rút viêm não ngựa Venezuela (VEE), Durden và cộng sự (1992) cho thấy, trong các thí nghiệm ở chuột thì D. gallinae là một véc tơ lan truyền vi rút theo cơ chế cơ học. Tuy nhiên, loài Laelaps kochi mang vi rút cũng cho thấy không gây nhiễm sang chuột khỏe mạnh. Vi rút không có khả năng tăng sinh hoặc lan truyền Trans-ovarian đã được chứng minh.
Vì vậy, đối với các hình thức của viêm não ngựa do togaviruses, vai trò của D. Gallinae được xem là véc tơ lan truyền vi rút này theo cơ chế cơ học mặc dù vai trò chính xác của nó trong dịch tễ học của bệnh lý này vẫn cần phải được xác định lại. Tuy nhiên, đối với các loài mạt, chưa đủ cơ sở để khẳng định.
KHẢ NĂNG LAN TRUYỀN VI KHUẨN
Một số nhà nghiên cứu đã nhấn mạnh vai trò của các loài mạt về khả năng lan truyền vi khuẩn. Trong số các loài chịu trách nhiệm về lan truyền bệnh từ động vật sang người, vi khuẩn Francisella tula rensis gây bệnh tularaemia, vi khuẩn này được lan truyền theo cơ chế cơ học do các loài mạt thuộc họ Dermanyssoidea giống như Tabanidae, Simuliidae hoặc Ixodidae ... (Timofeeva, 1964; Petrov, 1971; Zuevskii năm 1976; Algazin & Bogdanov, 1978;. Lysy và cộng sự, 1979). Tương tự như vậy, vi khuẩn Salmonella, Yersinia, Listeria, Pasteurella và Bacillus cũng được tìm thấy trên các loài mạt này nhưng lại không có bằng chứng chúng có khả năng lan truyền (Sturman, 1965; Poyarkov & Avchinnikov, 1966; Wilson & MacDonald, 1967; Grebenyuk và cộng sự ., 1972; Petrov, 1975;. Sotnikova và cộng sự, 1977; Romasheva & Uzdenov, 1979;. Zeman và cộng sự, 1982).
Dermanyssus gallinae đã được chứng minh là một véc tơ lan truyền xoắn khuẩn nhưng nó không phải là vật chứa xoắn khuẩn trong tự nhiên. Borrelia bugdorferi gây bệnh Lyme, bị nghi là đã được lan truyền bởi Ornithonyssus bacoti bởi một số nhà nghiên cứu (Ciolca và cộng sự, 1968;. Reshetnikov, 1967;. Stajkovic và cộng sự, 1993;. Lopatina và cộng sự, 1999). Cuối cùng, D. gallinae được cho là có khả năng lan truyền và vật chứa vi khuẩn Erysipelothrix rhusiopathiae, tác nhân gây bệnh viêm quần ở lợn (Chirico và cộng sự., 2003).
Bảng 3. Các loài mạt có khả năng lan truyền vi khuẩn
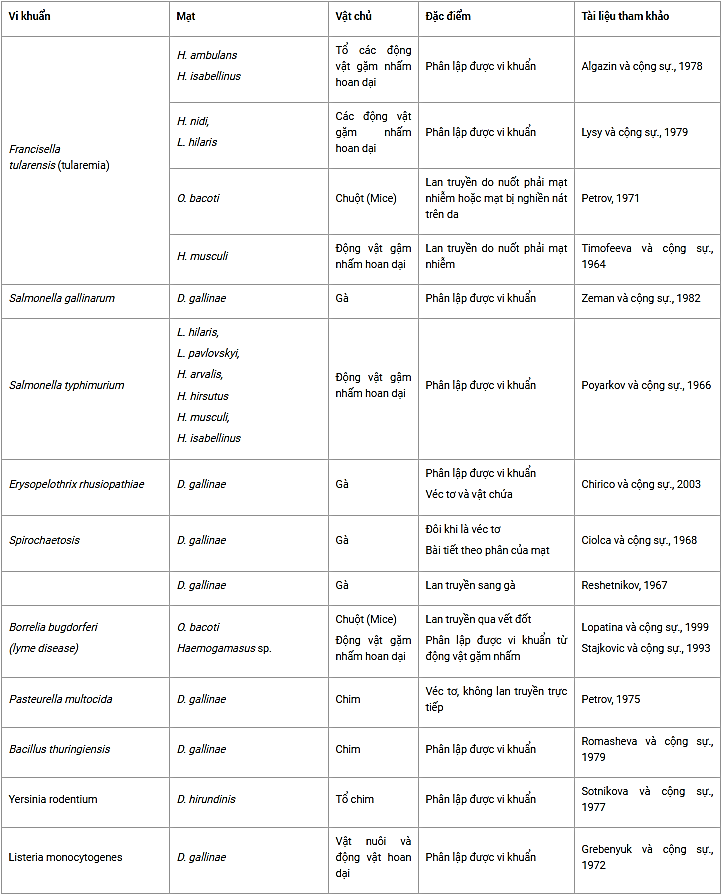
(D. hirundinus = Der- manyssus hirundinis, H. ambulans = Haemogamasus ambulans, H. arvalis = Hyperlaelaps arvalis, H. isabellinus = Hirstionyssus isabel- linus, H. hirsutus = Haemogamasus hirsutus, H. musculi = Hirstionyssus musculi, H. nidi = Haemogamasus nidi, L. hilaris = Laelaps hilaris, L. pavlovskyi = Laelaps pavlovskyi, O. bacoti = Ornithonyssus bacoti).
KHẢ NĂNG LAN TRUYỀN KÝ SINH TRÙNG
Một số nhà khoa học đã chứng minh vai trò của Ornithonyssus bacoti như một véc tơ lan truyền Trypanosoma cruzi theo cơ chế lan truyền cơ học trong điều kiện phòng thí nghiệm (Cortez-Jimenez, 1994) và đã thành công trong thử nghiệm về khả năng lan truyền cơ học đối với các đơn bào khác như Hepatozoon sp. và Elleipsisoma thomsoni của loài mạt này (Redington & Jachowski, 1971; Frank, 1977; Mohamed và cộng sự, 1987). Giun chỉ thuộc giống Litomosoides cũng đã được chứng minh là có khả năng phát triển ở bên trong các loài mạt thuộc họ Dermanyssoidea (Renz & Wenk, 1981; Diagne và cộng sự, 1989).
Bảng 4. Các loài mạt có khả năng lan truyền ký sinh trùng
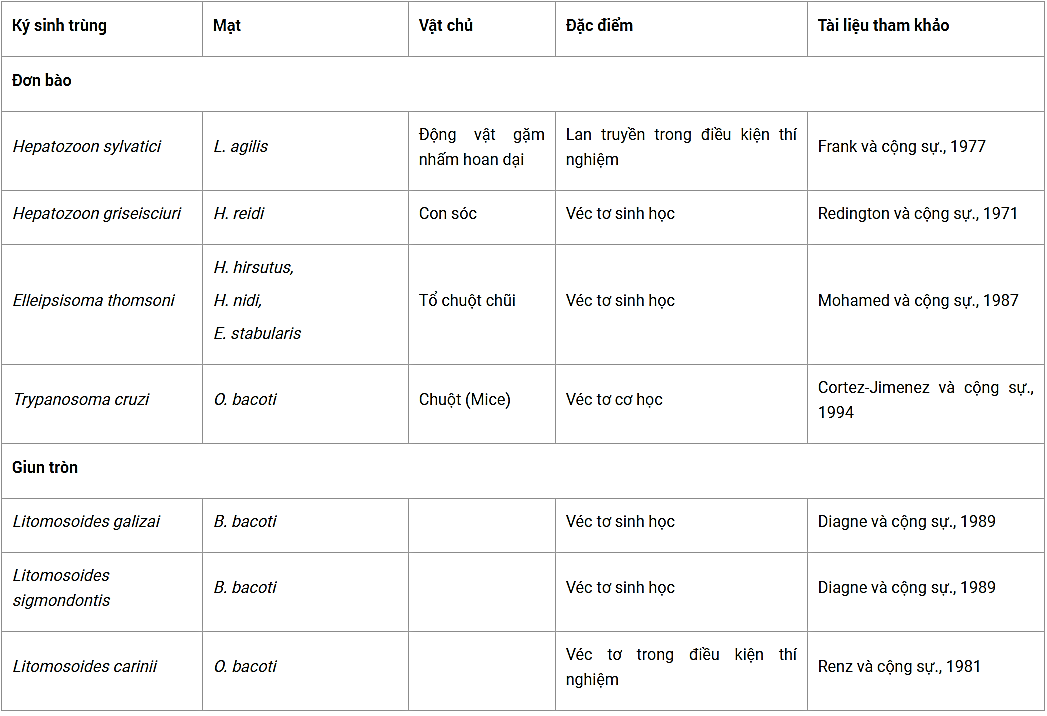
(B. bacoti = Bdellonyssus bacoti, E. stabularis = Eulaelaps stabularis, H. hirsutus = Haemogamasus hirsutus, H. nidi = Haemogamasus nidi, H. reidi = Haemogamasus reidi, L. agilis = Laelaps agilis, O. bacoti = Ornithonyssus bacoti).
KHẢ NĂNG LAN TRUYỀN RICKETTSIA
Các loài mạt thuộc phụ họ Dermanyssoidea có vai trò quan trọng trong việc lan truyền tác nhân gây bệnh là Rickettsia và Coxiella burnetii. Huebner và cộng sự (1946), là người đầu tiên nghĩ rằng Rickettsia akari được lan truyền bởi Liponyssoides sanguineus và loài mạt này cũng là vật chứa Rickettsia akari. Tuy nhiên, Yunker và cộng sự (1975) đã không phân lập được các loài Rickettsia khác từ các động vật gặm nhấm thường bị ký sinh bởi các loài mạt như Laelaps dearmasi, Ornithonyssus bacoti, Eubrachylaelaps jamesoni và Haemogamasus glasgowi mặc dù một số nhà khoa học, trích dẫn trong bài viết của họ, đã chứng minh sự tồn tại của rickettsia trong giống mạt Laelapidae một thời gian dài vào mùa đông, cũng như lan truyền theo cơ chế Trans-tadial, Trans-ovarian và lan truyền vật từ chủ này sang vật chủ khác của một số loài mạt thuộc họ Dermanyssoidea.
Rehacek và cộng sự (1975) đã tiến hành gây nhiễm rickettsia cho giống mạt thuộc họ Dermanyssoidea và cho thấy những loài mạt này đã tham gia vào quá trình lưu hành của các vi khuẩn và nó đã lan truyền vi khuẩn sang các động vật có vú nhỏ nhưng không thể kết luận vai trò lan truyền của chúng, có thể vai trò của chúng như là vật chứa hoặc véc tơ sinh học hoặc véc tơ cơ học. Ngược lại, Sonenshine và cộng sự (1978) đã đưa ra giả thuyết rằng cả hai loài mạt Haemogamasus reidi và Androlaelaps fahrenholzi đã tham gia vào việc lan truyền R. prowazekii sau khi quan sát một số lượng lớn các con sóc bị nhiễm tự nhiên nhưng phân tích huyết thanh học từ những động vật trong thời gian ủ bệnh do tiêm dịch huyền phù chứa Coxiella burnetii (người ta nghiền 9 loài mạt thuộc họ Dermanyssoidea bao gồm Allodermanyssus sanguineus và Hirstionyssus criceti mang Coxiella burnetii) kết quả không phát hiện được Coxiella burnetii, kể cả huyết thanh của các loài chim và động vật gặm nhấm, thu thập từ những nơi xảy ra dịch sốt Q (Zemskaya & Pchelkina, 1968).
Kocianova và cộng sự (1989, 1993) gây nhiễm rickettsia cho các loài mạt Haemogamasus nidi, H. hirsutus, Eulaelaps stabularis, Androlaelaps fahrenholzi và A. casalis sau đó đã tìm thấy vi khuẩn này trong phân của mạt.
Cuối cùng, Zemskaya & Pchelkina (1967) đã chứng minh bằng thực nghiệm rằng Dermanyssus gallinae, Ornithonyssus bacoti và Allodermanyssus sanguineus đều có khả năng mang rickettsia khi chúng đốt máu động vật bị nhiễm và vi khuẩn có thể tồn tại ít nhất 6 tháng ở mạt sống, và lên dến một năm ở mạt chết. Trong điều kiện thí nghiệm D. Gallinae đã lan truyền C. burnetii từ một con lợn guinea sang chim và ngược lại qua việc đốt máu của mạt. Loài O. bacoti cũng có thể lan truyền tác nhân gây bệnh giữa lợn guine và chim, giữa các con chim với nhau, và từ chim sang lợn guinea, lan truyền theo cơ chế Trans-ovarian cũng được ghi nhận. Tương tự như vậy, loài A. sanguineus có thể lan truyền rickettsia giữa các loài chim và động vật gặm nhấm (chuột và chuột lang).
Vì vậy, đối với vi khuẩn thuộc giống Rickettsia, chỉ có loài mạt Allodersanguineus manyssus thực sự được xem là véc tơ chính lan truyền R. akari. Đối với các loài rickettsia khác, một số được phân lập từ các loài mạt thuộc phụ họ Dermanyssoidea nhưng khả năng lan truyền của các loài mạt này cần được nghiên cứu. Cuối cùng, người ta đã được chứng minh rằng một số loài mạt thuộc họ Dermanyssoidea (O. bacoti, D. gallinae và A. sanguineus) không phải là véc tơ lan truyền C. burnetii trong điều kiện thí nghiệm và sự tồn tại lâu dài của các vi khuẩn trong mạt đã được quan sát. Lan truyền dọc của O. bacoti cũng đã được chứng minh (Zemskaya & Pchelkina, 1967).
Bảng 5. Các loài mạt có khả năng lan truyền rickettsia
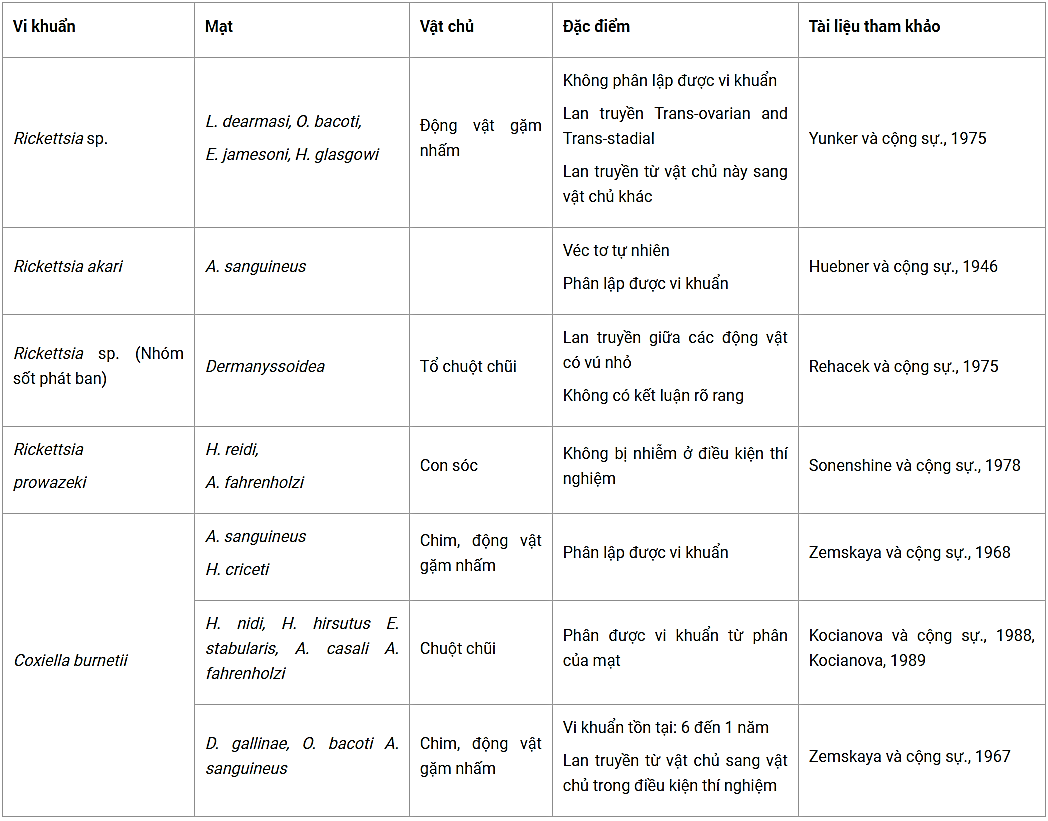
(A. casalis = Androlaelaps casalis, A. fahrenholzi = Androlaelaps fahrenholzi, A. sanguineus = Allodermanyssus sanguineus, E. jamesoni = Eubrachylaelaps jamesoni, E. stabularis = Eulaelaps stabularis, H. criceti = Haemogamasus criceti, H. glasgowi = Haemolaelaps glasgowi, H. hirsutus = Haemogamasus hirsutus, H. nidi = Haemoga- masus nidi, H. reidi = Haemogamasus reidi, L. dearmasi = Laelaps dearmasi, O. bacoti = Ornithonyssus bacoti).
NHẬN XÉT
Một số loài mạt rõ ràng là véc tơ lan truyền tác nhân gây bệnh, nhưng rất nhiều nghiên cứu cần phải thực hiện nếu chúng ta muốn hiểu được vai trò truyền bệnh của những loài mạt này. Từ quan điểm sinh học, chúng ta cần phải xác định chính xác các loài có liên quan đến các tác nhân gây bệnh.
Để làm điều này, cần phải xây dựng mô hình dựa trên một trong hai loài mạt Dermanyssus gallinae hoặc Ornithonyssus bacoti và lặp lại các nghiên cứu trước đây sử dụng các kỹ thuật sinh học phân tử hiện đại để phát hiện các tác nhân gây bệnh (Cortinas và cộng sự., 2002) và nghiên cứu chu kỳ của các loài mạt này trong điều kiện in vitro (Bruneau và cộng sự., 2001).
Vai trò của các véc tơ này trong dịch tễ học của các bệnh lan truyền từ động vật sang người trong mối quan hệ với véc tơ chính (đặc biệt là các loài ve) và với sự tồn tại liên tục và phổ biến của các tác nhân gây bệnh cần phải được xác định.
Bệnh viện Nguyễn Tri Phương - Đa khoa Hạng I Thành phố Hồ Chí Minh
BÀI VIẾT KHÁC


